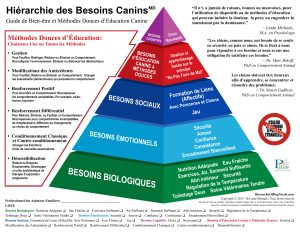
Quel impact de la domestication du chien sur leur alimentation ? Le régime alimentaire qui leur serait adapté est-il vraiment celui du loup ?
Des études récentes suggèrent que les chiens ont été domestiqués en Sibérie il y a environ 23 000 ans, alors que les loups et les hommes étaient isolés par le climat rigoureux du dernier maximum glaciaire. Les canidés ont ensuite accompagné les premiers hommes dans les Amériques, voyageant avec eux lors de leur dispersion il y a environ 15 000 ans (Perri et al., 2021). À notre connaissance, les chiens sont les premiers animaux domestiqués par l’homme et les seuls par les chasseurs-cueilleurs. Les hypothèses concernant la domestication comprennent la sélection par l’homme des loups les plus dociles et l’autodomestication par sélection naturelle des loups qui osaient se nourrir à proximité des campements humains. Un modèle basé sur des agents prédit que cette dernière hypothèse est la plus cohérente (Kulwicki et al., 2019), mais des incertitudes subsistent. En effet, le cloisonnement des niches écologiques des chiens, des loups et des renards commensaux individuels a été observé pour les canidés magdaléniens. Il a été démontré que les loups avaient un accès illimité et permanent aux ressources des espèces proies, tandis que les chiens voyaient leur régime alimentaire contrôlé par les humains (Baumann et al., 2020).
En tant que chasseurs, en meute, de grandes proies, dans des niches écologiques qui se chevauchent partiellement, les loups et les humains étaient en concurrence pour les ressources. Selon une étude récente, la répartition des ressources alimentaires entre les humains et les chiens naissants (en tant qu’espèce) pourrait avoir amélioré la compétition pendant la phase initiale de la domestication, permettant ainsi la coopération. En effet, la capacité du foie humain à métaboliser les protéines est limitée, ce qui nuit à la consommation de viande chez l’homme. Cette limitation aurait laissé un surplus de protéines d’origine animale disponible pour le partage avec les chiens naissants. Cette théorie repose sur le fait que les sites archéologiques du Pléistocène, où l’on trouve des restes de chiens naissants sont situés dans des régions où les conditions étaient alors analogues à celles des environnements arctiques ou subarctiques, qui induisaient que le gibier était dépourvu de graisse pendant les hivers rigoureux (Lahtinen et al., 2021).
Domestication du chien et alimentation : la digestion de l’amidon
Cette théorie est cohérente avec les résultats soulignant que l’adaptation des chiens à un régime riche en amidon n’a peut-être pas eu lieu lors de la domestication initiale, mais plutôt lorsque les chiens déjà domestiqués ont été confrontés à la révolution agricole, au cours du Néolithique. En effet, la sélection pour la digestion de l’amidon au cours de la domestication des chiens a ciblé le gène codant pour l’amylase pancréatique (AMY2B). Cette enzyme décompose l’amidon en maltose, et montre une multiplication par 5 en moyenne du nombre de copies du gène AMY2B et une activité plus élevée chez les chiens par rapport aux loups (Axelsson et al., 2013 ; Arendt et al., 2012, in Axelsson, 2018). Le nombre de copies du gène AMY2B indique la capacité à digérer l’amidon. La sélection chez le chien a également induit une expression accrue du gène codant pour la maltase-glucoamylase (MGAM), afin de convertir le maltose en glucose. Cela montre que la sélection a ciblé l’ensemble de la voie de digestion de l’amidon et l’absorption subséquente du glucose. Cependant, cette efficacité dans la digestion de l’amidon diffère selon l’origine des chiens. En effet, certains chiens n’ont pas de copies supplémentaires d’AMY2B, ce qui suggère qu’un changement de régime alimentaire a pu se produire chez des chiens déjà domestiqués, alors qu’ils suivaient les humains et leurs changements de régime et de mode de vie. En effet, cinq races de chiens se sont révélées relativement peu affectées par les adaptations métaboliques à la digestion de l’amidon : le Husky de Sibérie, le chien du Groenland, le chien de traîneau du Groenland, le Dingo, et peut-être Samoyède. Ainsi, les chiens originaires d’Australie, d’Amérique arctique et d’Asie arctique portent généralement moins de copies de MAY2B que les chiens originaires d’autres régions. Cette répartition est cohérente avec la diffusion de l’agriculture préhistorique, et son impact supposé sur le changement de régime alimentaire chez les chiens (Axelsson, 2018).
Cependant, une autre étude, utilisant une stratégie d’échantillonnage élargie, suggère que l’expansion du nombre de copies d’AMY2B pourrait avoir eu lieu avant la scission des grandes races modernes, les Huskies sibériens présentant une gamme de nombre de copies allant de 2 à 15 comme les autres races. Ainsi, la sélection positive a continué à influencer le nombre de copies d’AMY2B après l’expansion initiale, dans les races exposées à des régimes riches en amidon. En effet, la consommation d’amidon semble avoir influencé le nombre de copies d’AMY2B, la phylogénie n’expliquant pas ces nombres de copies. Les auteurs proposent donc que les régimes riches en amidon aient prédominé dans la plupart des races de chiens pendant ou peu après l’expansion et la prolifération de la variation du nombre de copies d’AMY2B, sous l’influence d’une pression de sélection positive due à une consommation élevée d’amidon. Ensuite, la migration et la différenciation des races à haute et basse teneur en amidon ont coïncidé avec la consommation locale d’amidon par les humains. La consommation élevée d’amidon a donc induit une pression sélective positive qui a entraîné une augmentation du nombre de copies d’AMY2B. La pression de sélection s’est ensuite maintenue chez les chiens consommant des régimes enrichis en amidon, et la variation du nombre de copies d’AMY2B a fait l’objet d’une dérive génétique chez les chiens ayant adopté des régimes pauvres en amidon, augmentant la variation dans l’ensemble de la race (Reiter et al., 2016).
Ces études montrent l’importance de prendre en compte la domestication du chien dans l’alimentation, notamment la race ou le croisement particulier du chien avec lequel on travaille, afin de proposer une solution adaptée. En effet, une alimentation sans céréales peut être intéressante pour certaines races comme les huskies, surtout si des problèmes digestifs peuvent être à l’origine de problèmes de santé ou de comportement. J’ai constaté moi-même, à travers mon expérience personnelle de plus de 15 ans avec mes propres huskies sibériens, bergers, et chiens de connaissances, que le métabolisme et le goût des huskies sibériens semblent différents : ils semblent plus friands de sources alimentaires grasses, et moins ouverts à toute source alimentaire telle que certains légumes. De plus, les effets attendus des aliments industriels pour chiens ne sont parfois pas observés, et des effets contraires peuvent même être constatés.
Domestication du chien et alimentation : effets sur le microbiote
Par ailleurs, il est bien connu que le microbiote joue un rôle important dans la digestion. Chaque individu ayant son propre microbiote, la digestion diffère potentiellement d’un chien à l’autre en raison des cellules qui colonisent leur tube digestif. Le milieu de vie des chiens a permis l’adaptation de leur microbiote à une alimentation riche en amidon (Liu et al., 2021). En effet, des études du microbiome intestinal de coprolithes canins datés de 3600 à 3450 ans ont révélé que la structure de leur microbiome se trouvait dans une sorte d’état de transition entre celui des chiens modernes et celui des loups sauvages. La niche alimentaire déduite était omnivore, avec une consommation d’aliments agricoles riches en amidon. En outre, le microbiome s’est révélé enrichi en séquences codant pour des alpha-amylases, en complément d’un faible nombre de copies du gène de l’amylase de l’hôte (AMY2B). Ainsi, la transition vers un régime riche en amidon aurait pu être possible grâce à l’expansion du catabolisme microbien de l’amidon, avant une réponse génétique retardée de l’hôte aux nouvelles ressources alimentaires abondantes en amidon (Rampelli et al., 2021).
En effet, les effets de l’alimentation sur la santé sont à la fois primordiaux et subtils, et les effets observés ne sont pas toujours ceux attendus. Par exemple, un régime sans céréales contenant les plus grandes quantités d’amidon résistant à la digestion, d’amidon lentement digestible, et les plus faibles quantités d’amidon rapidement digestible et d’amidon total, a induit une vidange gastrique plus importante et plus rapide chez les Huskies par rapport aux régimes à base de céréales complètes, traditionnels et végétaliens, et un contrôle du glucose. Les auteurs ne s’attendaient pas à ce résultat, car un amidon plus résistant était censé induire une glycémie postprandiale plus faible et des valeurs d’indice glycémique plus basses. Les chercheurs ont donc émis l’hypothèse que les différences de macronutriments entre les aliments testés pouvaient avoir influencé les résultats (Richards et al., 2021).
Cependant, le changement microbien peut n’être que temporaire en cas de changement de régime alimentaire. En effet, l’analyse de la population microbienne intestinale des chiens lors du passage d’un régime de croquettes transformées riches en amidon à un régime de viande crue proche de la nature, en utilisant les loups comme référence, a montré que certains groupes microbiens se rapprochaient du profil microbien des loups, tandis que d’autres ne présentaient que des changements temporaires lors des changements d’alimentation. Les bactéries fermentant les glucides ont cédé la place aux fermenteurs de protéines lorsque les chiens sont passés d’un régime de croquettes à un régime cru, et certaines populations du microbiote intestinal se sont rapprochées de celui des loups. Les différences entre les loups et les chiens subsistent après le changement de régime (Xu et al., 2021).
Chez les chiens, certains taxons du microbiote liés au métabolisme des glucides ont été observés en plus grand nombre que chez les loups. Une différence significative a été mesurée dans les gènes codant pour la famille 34 des glycosyltransférases, la famille 25 des modules de liaison aux hydrates de carbone et la famille 13 des glycosides hydrolases, entre les métagénomes du microbiote intestinal des loups gris et des chiens domestiques. Le microbiote intestinal des chiens domestiques a également montré une plus grande biosynthèse de leucine, d’isoleucine et de valine et un plus grand métabolisme de l’azote, ce qui montre que le régime alimentaire des chiens domestiques contient moins de protéines que celui des loups. La fonction et l’abondance du microbiote intestinal des chiens domestiques se sont adaptées à la domestication (Lyu et al., 2018). La sélection artificielle et la cohabitation des chiens avec leurs propriétaires influencent le microbiote de l’intestin canin par l’acquisition et la perte de taxons bactériens, et la composition bactérienne et le métabolisme du microbiote de l’intestin canin se sont adaptés à un apport élevé en glucides ou en protéines. Ainsi, la domestication et la sélection ont affecté à la fois le génome canin et la population bactérienne de l’intestin canin (Alessandri et al., 2019).
Métabolisme
D’autres analyses génomiques de chiens et de loups ont permis d’observer des différences métaboliques. L’analyse des variations structurelles du génome du chien a montré un enrichissement du métabolisme énergétique, avec une insertion au niveau de la population d’une copie du transcrit Aldo-Keto Reductase Family Member B (AKR1B1). Cette nouvelle copie d’AKR1B1 est fortement exprimée dans l’intestin grêle et le foie, induisant une augmentation de la capacité antioxydante et des synthèses de novo d’acides gras chez le chien par rapport au loup gris, ce qui est probablement une réponse aux changements de régime alimentaire au cours de la révolution agricole. Ainsi, la rétroposition, qui est l’insertion de fragments d’ADN dans les chromosomes après leur transcription à partir de l’ARNm, peut impliquer de nouveaux gènes, facilitant ainsi la domestication (Wang et al., 2019).
Domestication du chien et alimentation : Conclusion
L’alimentation est un aspect important de la vie d’un chien. En effet, la thérapie métabolique peut être développée pour traiter certaines maladies, comme l’épilepsie. Par exemple, il a été montré qu’un régime alimentaire avec une teneur réduite en glucides permettait de contrôler les crises, par rapport à la prise de médicaments antiépileptiques (Masino et al., 2018). C’est pourquoi, comprendre l’importance de la diversité, à la fois dans le génome du chien, et dans son microbiote peut aider à mettre en place des solutions adaptées et personnalisées pour résoudre les problèmes des chiens actuels. D’après ces études, la domestication du chien a impacté son alimentation, et son métabolisme ainsi que son microbiote.
Besoin d’aide avec votre chien ?

Contactez-moi ! Comportementaliste et éducatrice canin diplômée, j’interviens en Lorraine, Vosges, ainsi qu’à distance en visio.
Sources
Alessandri, G., Milani, C., Mancabelli, L., Mangifesta, M., Lugli, G. A., Viappiani, A., Duranti, S., Turroni, F., Ossiprandi, M. C., van Sinderen, D., & Ventura, M. (2019). Metagenomic dissection of the canine gut microbiota: insights into taxonomic, metabolic and nutritional features. Environmental Microbiology, 21(4), 1331–1343. https://doi.org/10.1111/1462-2920.14540
Axelsson, E. (2018). Adapting to a new diet during dog domestication–how dogs coped with the agricultural revolution. Hill’s Global Symposium, 3–6.
Baumann, C., Starkovich, B. M., Drucker, D. G., Münzel, S. C., Conard, N. J., & Bocherens, H. (2020). Dietary niche partitioning among Magdalenian canids in southwestern Germany and Switzerland. Quaternary Science Reviews, 227, 106032. https://doi.org/10.1016/j.quascirev.2019.106032
Kulwicki, R., & Capaldi, A. (2019). Wolf Domestication: An Agent-based Simulation. Symposium on Undergraduate Research and Creative Expression.
Lahtinen, M., Clinnick, D., Mannermaa, K., Salonen, J. S., & Viranta, S. (2021). Excess protein enabled dog domestication during severe Ice Age winters. Scientific Reports, 11(1). https://doi.org/10.1038/s41598-020-78214-4
Liu, Y., Liu, B., Liu, C., Hu, Y., Liu, C., Li, X., Li, X., Zhang, X., Irwin, D. M., Wu, Z., Chen, Z., Jin, Q., & Zhang, S. (2021). Differences in the gut microbiomes of dogs and wolves: roles of antibiotics and starch. BMC Veterinary Research, 17(1). https://doi.org/10.1186/s12917-021-02815-y
Lyu, T., Liu, G., Zhang, H., Wang, L., Zhou, S., Dou, H., Pang, B., Sha, W., & Zhang, H. (2018). Changes in feeding habits promoted the differentiation of the composition and function of gut microbiotas between domestic dogs (Canis lupus familiaris) and gray wolves (Canis lupus). AMB Express, 8(1). https://doi.org/10.1186/s13568-018-0652-x
Masino, S. A., Freedgood, N. R., Reichert, H. R., Director, C. J., Whittemore, V. H., & Zupec-Kania, B. (2019). Dietary intervention for canine epilepsy: Two case reports. Epilepsia Open, 4(1), 193–199. https://doi.org/10.1002/epi4.12305
Perri, A. R., Feuerborn, T. R., Frantz, L. A. F., Larson, G., Malhi, R. S., Meltzer, D. J., & Witt, K. E. (2021). Dog domestication and the dual dispersal of people and dogs into the Americas. Proceedings of the National Academy of Sciences, 118(6). https://doi.org/10.1073/pnas.2010083118
Rampelli, S., Turroni, S., Debandi, F., Alberdi, A., Schnorr, S. L., Hofman, C., Taddia, A., Helg, R., Biagi, E., Brigidi, P., D&Apos;Amico, F., Cattani, M., & Candela, M. (2021). Bronze Age Coprolites Unveil the Buffering Role of the Gut Microbiota in the Dietary Adaptation of Dogs to Domestication. SSRN Electronic Journal. https://doi.org/10.2139/ssrn.3850374
Reiter, T., Jagoda, E., & Capellini, T. D. (2016). Dietary Variation and Evolution of Gene Copy Number among Dog Breeds. PLOS ONE, 11(2), e0148899. https://doi.org/10.1371/journal.pone.0148899
Richards, T. L., Rankovic, A., Cant, J. P., Shoveller, A. K., Adolphe, J. L., Ramdath, D., & Verbrugghe, A. (2021). Effect of Total Starch and Resistant Starch in Commercial Extruded Dog Foods on Gastric Emptying in Siberian Huskies. Animals, 11(10), 2928. https://doi.org/10.3390/ani11102928
Wang, G. D., Shao, X. J., Bai, B., Wang, J., Wang, X., Cao, X., Liu, Y. H., Wang, X., Yin, T. T., Zhang, S. J., Lu, Y., Wang, Z., Wang, L., Zhao, W., Zhang, B., Ruan, J., & Zhang, Y. P. (2018). Structural variation during dog domestication: insights from gray wolf and dhole genomes. National Science Review, 6(1), 110–122. https://doi.org/10.1093/nsr/nwy076
WOJTAŚ, J., KARPIŃSKI, M., TAJCHMAN, K., CZYŻOWSKI, P., GOLEMAN, M., ZIELIŃSKI, D., KOZAK, A., KRUPA, W., GARBIEC, A., & DROZD, L. (2018). Canis lupus familiaris – DOMESTICATION. Folia Pomeranae Universitatis Technologiae Stetinensis Agricultura, Alimentaria, Piscaria et Zootechnica, 345(48), 129–136. https://doi.org/10.21005/aapz2018.48.4.13
Xu, J., Becker, A. A. M. J., Luo, Y., Zhang, W., Ge, B., Leng, C., Wang, G., Ding, L., Wang, J., Fu, X., & Janssens, G. P. J. (2021). The Fecal Microbiota of Dogs Switching to a Raw Diet Only Partially Converges to That of Wolves. Frontiers in Microbiology, 12. https://doi.org/10.3389/fmicb.2021.701439